KASVINJALOSTUS / Matti Pekkarinen
1. Perinnöllisyyden perusteet
(perinnöllisyys ilmenee siten, että yksilö muistuttaa ominaisuuksiltaan vanhempiaan enemmän kuin lajin muita yksilöitä ... ja tietenkin sitenkin, että tietyn lajin jälkeläiset ovat samaa lajia)
a) perimä ja ilmiasu
Yksilön ominaisuudet (ulkonäkö ym) määräytyvät perimän ja ympäristön yhteisvaikutuksena. "ilmiasu=perimä+ympäristö", eli fenotyyppi=genotyyppi+ympäristö.
b) muuntelu
Kunkin lajin sisällä on erilaisia yksilöitä, myös perimältään. Aikojen kuluessa on kehittynyt mm. ympäristötekijöiden valintapaineen vuoksi erilaisia "selviytymistapoja", uusia ominaisuuksia. Lapin kuuset ovat riippuoksaisia (pärjää lumikuorman kanssa paremmin kuin eteläinen tyyppi, tosin menettää osan auringonvalosta epäedullisen oksa-asennon vuoksi.)
c) perintötekijät ovat kromosomeissa
Kaikkien solujen tumassa on lajille tyypillinen määrä kromosomeja. Ne ovat pitkiä DNA-rihmoja. DNA on eräs ns. nukleiinihappo. Se on molekyylirakenteeltaan tikapuun kaltainen. Pystyrunkona vuorottelevat fosforihappotähde ja sokeri (deoksiriboosi) ja poikkipuina emäsparit. Emäksiä on neljä erilaista (adeniini, tymiini, guaniini ja sytosiini, eli A,T,G,C). Kaikkia kromosomeja on pareittain (molemmilta vanhemmilta yksi). Niinpä kutakin geeniä on myös kaksi. Geeni on "pätkä kromosomia".
Jokainen kolmen peräkkäisen emäksen sarja kromosomissa määrää kasvin proteiinisynteesissä tietyn aminohapon. Noin sadan tällaisen "tripletin" sarja muodostaa geenin. Täten, ohjaamalla aminohappojen tuotantoa eliön soluissa, geenit vaikuttavat ja määräävät eliön rakennetta ja ominaisuuksia.
d) lokus
Kullakin geenillä on tietty sijainti: juuri tietyssä kromosomissa ja siinäkin tietyssä paikassa. Tutkimus on jo pystynyt selvittämään joidenkin eliöiden geenikartan. Geenit sijaitsevat pääasiallisesti kromosomien DNA:ssa, joskin myös mitokondrioiden ja plastidien rengasmaisesta DNA:sta on löydetty geenejä.
e) alleelit
Kustakin geenistä esiintyy useita erilaisia alleeleja, "versioita" (esim ihmisen silmän väri, veriryhmät jne)
f) DNA JA VALKUAISAINESYNTEESI
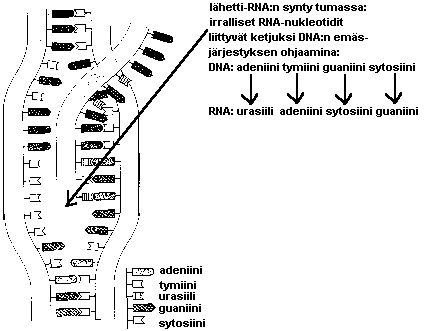
Kun jokin geeni alkaa toimia, DNA:n kaksoiskierre
purkautuu sen kohdalta ja sen emäsjärjestys kopioituu lähetti-RNA:n muotoon.
Lähetti-RNA vie geenin "viestin" ulos tumasta solulimassa oleviin ribosomeihin, joissa valkuaisaineet muodostuvat.
Siirtäjä-RNA:t tuovat ribosomille aminohappoja ja tunnistavat lähetti-RNA:sta kohdan, johon kukin aminohappo muodostuvassa valkuaisaineessa tulee. Näin DNA:n (geenien) emäsjärjestys muuttuu tietyksi aminohappojärjestykseksi valkuaisaineessa.
Syntynyt valkuaisaine
(entsyymi) puolestaan säätelee ja ohjaa tiettyä reaktiota solussa. Ominaisuus
muodostuu vasta, kun solujen ja kudosten erilaistumiseen kuuluvat
entsyymireaktiot tapahtuvat. Sen vuoksi geenien ilmentymisen eli ekspression säätely on perimmäinen syy solujen erilaisuudelle.
Tämä lähetti-RNA:n synteesi tapahtuu tumassa... sieltä valmis lähetti-RNA siirtyy tumakelmun aukkojen kautta solulimaan, jossa proteiinisynteesi tapahtuu:
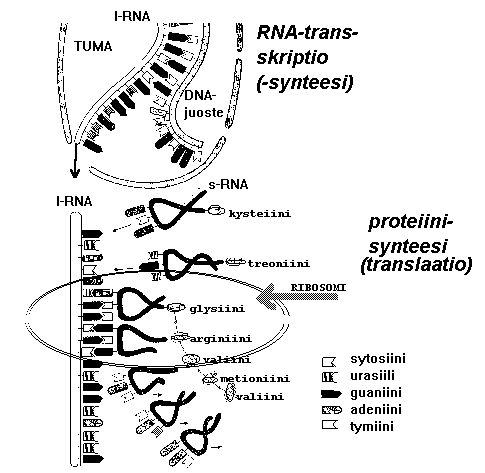
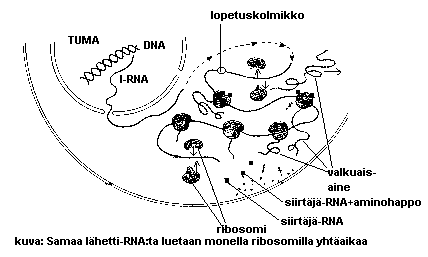
Yl. soluissa vain osa entsyymejä tuottavista ns.
rakennegeeneistä on toiminnassa. Eri geenit ovat aktiivisia yksilönkehityksen
eri vaiheissa. Juuri tämä mahdollistaa sen solujen ja kudosten erilaistumisen,
joka on monisoluisten eliöiden yksilönkehityksen ja elintoimintojen perusta.
Ns. pleiotrooppiset geenit vaikuttavat useaan ominaisuuteen. Polygeenit taas
vaikuttavat erillisyydestään huolimatta samaan ominaisuuteen ja säätelevät
usein ns. kvantitatiivisia ominaisuuksia, esim. kasvua. Letaaligeenit
aiheuttavat yksilön kuoleman.
g) tumanjakautuminen
= solunjakautumista edeltävä tapahtumaketju, jota on kaksi tyyppiä: mitoosi, jossa syntyneet tytärsolut sisältävät täsmälleen samat kromosomit kuin emosolukin, ja meioosi, jossa tytärtumien kromosomien lukumäärä vähenee puoleen.
Meioosissa "arpoutuvat" kromosomit syntyvään sukusoluun (joko äidiltä tai isältä saatu kromosomi päätyy siihen kustakin (ihmisellä 23) kromosomiparista.
Mitoosissa kromosomit halkeavat kahdeksi identtiseksi DNA-rihmaksi. Perimä ei muutu.
Mitoosin esivaiheessa, profaasissa,
pitkittäin kahdentuneet kromosomit alkavat erottua kromoneemarihmojen
spiralisoitumisen takia, tumajyvänen ja -kelmu häviävät. Samanaikaisesti on
sentrosomi jakautunut kahtia, ja sen puoliskot siirtyvät tuman vastakkaisille
puolille. Tumasukkula alkaa muodostua niiden väliin. Keskivaiheessa,
metafaasissa, yhä selvemmin näkyvät kromosomit siirtyvät vähitellen
sukkulan keskitasoon ja muodostavat metafaasilevyn. Ne halkeavat pituussuunnassa
kahdeksi samanlaiseksi tytärkromosomiksi. Jälkivaiheessa, anafaasissa, tytärkromosomit
eroavat ja kulkevat sukkulan päihin. Loppuvaiheessa, telofaasissa, tumasukkula
häviää, kromosomien kierteisyys purkautuu, ja ne häviävät näkyvistä.
Kummankin kromosomiryhmän ympärille muodostuu tumakelmu, ja tumajyväset
tulevat näkyviin. Tumasta on tullut kaksi identtistä tumaa. Mitoosin kestoaika
vaihtelee 10 minuutista useihin tunteihin. Jakautumisten välisenä aikana,
interfaasissa, kromosomit kaksinkertaistuvat seuraavaa jakautumisvaihetta
varten.
Meioosin eli kypsymisjakautumisen tuloksena syntyy lisääntymissoluja.
Siihen kuuluu kaksi perättäistä tumanjakautumista. Ensimmäinen on vähennys-
eli reduktiojakautuminen, jossa kromosomisto puolittuu. Toinen on tasaus- eli
ekvaatiojakautuminen eli tavallinen mitoosi.
Solujen jakautuessa mitoosissa (tumanjakautuminen) tulee tytärsoluihin samat geenit kuin emosolussakin oli. Sukupuolisoluihin, jotka muodostuvat meioosissa, tulee kutakin kromosomia (ja geeniä) vain yksi kappale. Sukupuolisolujen yhtyessä muodostuva uusi yksilö saa näin ominaisuutensa puoliksi kummaltakin vanhemmaltaan.
h) Perimän siirtyminen risteytyksessä: Monohybridiristeytys, jossa tarkastellaan YHDEN ominaisuuden periytymistä keskenään erilaisilla vanhemmilla. Mendelin esimerkki: vihreäsiemeninen herne X keltasiemeninen (P-polvi). KAIKKI jälkeläiset keltaisia (keltasiemenisyys on dominoiva eli vallitseva ominaisuus) =F1-sukupolvi. Seuraavassa polvessa (F2) on 1/4 vihreitä ja 3/4 keltaisia. Dihybridiristeytyksessä tarkkaillaan kahden ominaisuuden periytymistä niiden suhteen erilaisilla vanhemmilla. (hybridi tarkoittaa perinnöllisesti toisistaan poikkeavien vanhempien jälkeläistä, mm. eläinten roturisteytykset)
i) Ominaisuuksien (ja geenien) kytkentä. Samassa kromosomissa olevien geenien määräämät ominaisuudet eivät "mendelöi". Esim silmän väri ja ihon ja hiusten väri ovat ihmisillä toisiinsa kytkeytyneet. Tekijäinvaihdunta (crossing over): Kromosomiparin vastinkromosomien DNA-rihmat menevät meioosissa ristikkäin ja katkeavat. Tämä mekanismi purkaa geenien kytkennän.
j) Mutaatio: yksilön tai solun perimän äkillinen täysin sattumanvarainen muutos
mutaatio: yksilön tai solun perimän äkillinen täysin
sattumanvarainen muutos. Geenimutaatiossa yksittäisen geenin DNA:n emäsjärjestys
muuttuu esim. virheellisen replikaation takia. Muutos koskee yl. yhtä
nukleotidiparia (pistemutaatiot), ja yhdenkin geenin alueella niitä voi sattua
satoja erilaisia. Tällainen geenissä tapahtuva vähäinen muutos voidaan
todeta vain sen aiheuttaman ominaisuuden mahdollisesta muuttumisesta.
Kromosomimutaatiot ovat kromosomin katkeamisesta ja osien
uudelleenryhmittymisestä aiheutuvia rakennemuutoksia, jotka näkyvät usein
mikroskoopillakin. Kromosomista voi irrota ja hävitä pala joko päästä (pilkkouma,
fragmentaatio) tai keskeltä (häviämä, deleetio). Jokin kromosomin osa voi
kaksinkertaistua (kahdentuma, duplikaatio) tai kääntyä toisinpäin (kääntymä,
inversio). Kaksi ei-homologista kromosomia voi vaihtaa osia (siirtymä,
translokaatio). Kromosomistomutaatiossa koko genomi muuttuu. Aneuploidiassa on
yksi tai useampi ylimääräinen kromosomi tai kromosomeja puuttuu; trisomiassa
on diploidisella yksilöllä yksi kromosomi kolminkertaisena, monosomiassa
yksinkertaisena. Koko kromosomisto on moninkertaistunut polyploidiassa.
Somaattiset (eli muussa kuin lisääntymissolussa tapahtuvat) mutaatiot
aiheuttavat kasveille harhamia, eläimille mosaiikkikudoksia.
Mutantti on yksilö, jolla on mutaation seurauksena
normaalityypistä poikkeava perimä. Mutageeni on tekijä, joka lisää
mutaatioiden määrää. Mutageeneja ovat mm. ionisoiva ja lyhytaaltoinen säteily
(röntgen-, ultraviolettisäteily) sekä erilaiset nukleiinihappoihin
vaikuttavat yhdisteet (typpihapoke, monet ympäristömyrkyt).
2. Kasvien lisääntymistavat
2.1. Vegetatiivinen lisääntyminen
lisääntyminen
kasvullista, perinnöllistä
emoyksilöstä
monistamalla
...perinnölliset
ominaisuudet säilyvät rajattomasti
...heteroosi-ilmiön hyväksikäyttö, kaikki lajikkeet
Juolavehnä, peltovalvatti, nokkonen: lisääntyminen juurakon silmuista.... Marjapensailla lisääminen oksapistokkaista...
2.2. Suvuton
lisääntyminen
lisääntymiselimenä siemen, joka on syntynyt ilman
.yleistä
esim. FESTUCA-, POA- , CITRUS- ja RUBUS-suvuilla
-ilmiö ei
ole ehdoton
-jalostusmahdollisuudet
vähenevät koska meioosin
2.3 .
Suvullinen lisääntyminen
lisääntyminen kukan avulla, hedelmöityksen kautta
vain suvullinen lisääntyminen aiheuttaa perinnöllistä muuntelua
2.3.1. Itsesiitos vai ristisiitos ?
kumpaa
menetelmää kasvilaji käyttää, on suhteellinen kysymys
ilmiö
riippuu siitä , kuinka tehokkaasti
kasvi pystyy estämään itse- tai sukulaispölytyksen
.on ns.
ITSEINKOMPATIBILITEETTI-faktori, esim. apilalla estää siiteputken kasvu emin
luotilla
2.3.2. Itsesiitos l. autogamia
kasvi on itsefertiili
kukan
rakenne estää ristisiitoksen, joskus mahdollista
lajeja:
ohra, vehnä, kaura ja herne
jalostus ja
lisäysviljely helppoa koska eristysvaatimukset ovat vähäisiä
geneettistä
taantumisilmiötä esim. ohralla ei voi tapahtua
2.3.3.
kukan rakenne pyrkii estämään itsepölytyksen
.estämisratkaisuja:
.....kukan
morfologia
.....hede- ja
emikukkien eriaikainen kehittyminen
..siitepöly
ei idä emin luotilla
jalostus ja siementen lisäysviljely
vaikeaa eristys vaatimuksen vuoksi
huomaa: ristisiittoisilla lajeilla kuten rukiilla,
timoteillä, apilalla ym. jalostuksessa
Koirassteriliteetti
-pyritään estämään populaatiossa
väärä pölytys
-on käytetty mm. hybridi-vehnän jalostuksessa
-mekaaninen tai kemiallinen keino
-heteet ( tai hedekukka) kerätään pois
-siitepölyn kehitys estetään
-MS-faktorin käyttö (male sterility, koirassteriliteetti)
-tervettä siitepölyä ei muodostu, geneettinen
-käyttö yleistä hybridilajikkeiden tuottamisessa
-periaate: pitkälle sukusiitettyjen linjojen välisissä rlsteytyksissä
saadaan
aikaan voimakas
heteroosivaikutus
-maissin tuotanto tapahtuu pääasiassa hybridisiemenellä
3. Jalostusmenetelmät
3.1. Valintajalostus
-työskentely
spontaanin muuntelun varassa, erillisenä jalostusmenetelmänä luovuttu koska
-metsänjalostuksessa PLUS-puut ja PLUS-metsiköt:valintahyötyä
on todettu
(-esim perunan lajittelu ei ole perimän jalostamista
(klooni, ei muuntelua)... entä rukiin lajittelu?)
3.2. Risteytysjalostus
-eniten käytetty
menetelmä
(nykyisin
jalostajat käyttävät useita menetelmiä yhdessä )
-risteyttämällä
saadaan aikaan laajaa muuntelua
-myös lajien
väliset risteytykset mahdollisia läheisten sukulaislajien välillä, esim.
ruisvehnä
-risteytysvanhempien
valinta tärkeä (muuntelun rajat)
-risteytyksen
jälkeen saadaan F1 -polvi, jossa heteroosi-ilmiö esiintyy voimakkaana
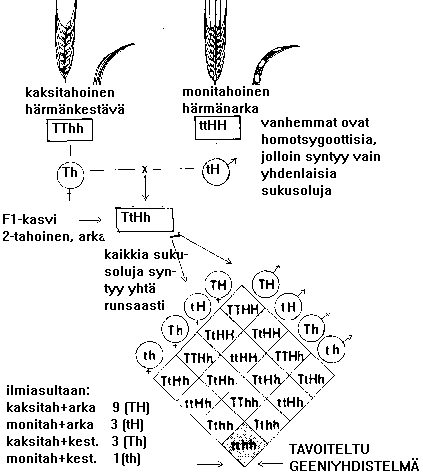
3.2.1. Itsesiittoiset kasvit
-aloitetaan
emaskuloinnilla, heteiden poisto pinseteillä
-ajankohdan arviointi: pölytys tapahtuisi 2 - 4 pv:n kuluttua
-pergamiini-pussi tähkän päälle , estetään kuivuminen ja vieraan
siitepölyn pääsy
Kuva: vehnän emaskulointi

-pölyttäjältä
tuodaan 2 - 3 pv: n kuluttua isäkasvin, heteen ponnet
-valinta yl.
F2-polvesta (valitaan tähkiä silmävaraisesti)
"pedigree-menetelmä"
-valintoja
F3- ja F4-Polvissa jatketaan, heikot pois
-jalostajilla
on 'omia' menetelmiään, jotkut aloittavat vasta F4- tai F5-Polvista (bulk-menetelmä)
3.2.2. Ristisiittoiset kasvit
-risteytysvanhemmat heterotsygoottisia ,
-valinta
mahdollista jo Fl-polvessa
menetelmiä:
1 . Dialleeliristeytys
-käytetään hyväksi 'periyttämiskykyä'
- testataan ensin mitkä vanhemmat sopivat
parhaiten toisilleen
2. Polycross-menetelmä
-muodostetaan
sekakasvustoja useista lajikkeista,
-sekoitustapoja
on useita, esim. ristiinkylvö
-valitaan
eristyksiin lisättäväksi haluttuja kombinaatioita
-laajojen
aineistojen hallinta ristisiittoisilla lajeilla on vaikeaa
3.3. Polyploidijalostus
-käytetään
hyväksi kromosomien määrän poikkeamista normaalista
-tavallisimpia
ovat peruskromosomiston ( 2n) kaksinkertaistumat (4n = tetraploidit) ja
kolminkertaistumat (6n = heksaploidit)
-saadaan
aikaan kolkisiinikäsittelyllä
-ovat
mutaatioita - ei kuulu mutaatiojalostukseen ?
3.4. Mutaatiojalostus
-pyritään
keinollisesti aikaansaamaan muutoksia perimässä (kromosomeissa, geeneissä)
-tavallisimpia
mutageenejä :
-fysikaaliset: gamma - ja rön-tgen säteilytys
-.kemialliset : etylmetansulfonaatti (EMS )
-mutantit
ovat harvoin suoraan käyttökelpoisia , käytetään yleisesti risteytyksiin
-esimerkkejä:
Ryhti, Puhti ja Nasta ovat ns. epäsuoria mutaatiojalosteita
3.5. Resistenssijalostus
-jalostusta
kasvien tauteja ja tuholaisia vastaan
-käytetään
hyväksi perinnöllisesti heterogeenistä aineistoa, jota stressataan esim.
taudeilla
.valitaan
parhaiten selviytyneitä yksilöitä jatkojalostukseen
Resistenssityypit :
1 .
pystysuora l. vertikaaliresistenssi
-isäntä/loinen
; geeni-geeniltä periaate
-pyritään estämään täysin tuhoojan hyökkäykset
-murtuu käytännössä
helposti
2.
vaakasuora 1. horisontaa1iresistenssi
=kenttäkestävyys
-on useiden
geenien aiheuttama ominaisuus
-jalostuksessa
haluttu ominaisuus nykyisin
-isäntä/loissuhteen tasapaino
3.6. Heteroosijalostus
3.7. Ylläpitojalostus
3.8. Protoplastifuusio
protoplasti, kasvisolu, jonka soluseinä on poistettu entsymaattisesti.
Protoplasti on herkkä ympäristön osmoottisille muutoksille ja mekaaniselle käsittelylle.
Suotuisissa oloissa protoplastit rakentavat soluseinän uudelleen eli
regeneroituvat normaaleiksi soluiksi. Eri kasvien protoplasteja voidaan
fuusioida keskenään kalsiumionien, polyetyleeniglykolin tai sähkökentän
avulla, jolloin muodostuu somaattisia hybridisoluja. Fuusioituneet solut voidaan
regeneroida kasveiksi, jolloin saadaan uudenlaisia risteymiä tai uusia
ominaisuuksia omaavia hyötykasveja.
4.Biotekniikka
Biotekniikka on laaja käsite
biotekniikka (< kreik.), luonnontieteiden ja insinööritieteiden
yhdistämistä tuotteiden ja palvelujen aikaansaamiseksi eliöiden, solujen,
niiden osien ja niissä esiintyvien molekyylien avulla. Biotekniikan perinteisiä
muotoja ovat mikrobien aiheuttamiin käymisreaktioihin perustuvat oluen ja
viinien valmistus, leivän nostattaminen hiivalla sekä juustojen, piimän,
hapankaalin yms. valmistus. Moderni biotekniikka hyödyntää geenitekniikan
(geenimanipulaatio) ja molekyyligenetiikan (molekyylibiologia) avulla soluja ja
niissä esiintyviä kem. reaktioita. Geenitekniikan
avulla geenejä voidaan siirtää mikrobista toiseen, mutta myös korkeammilla
eliöillä sekä yli lajirajojen, esim. ihmisestä mikrobiin tai kasviin.
molekyylibiologia, biologian osa, joka tutkii
elämän ilmiöitä molekyylitasolla, ts. selvittelee minkälaisiin
molekyyleihin eliöiden rakenteet ja toiminnot pohjautuvat. Molekyylibiologia
tutkii etenkin suuria nukleiinihappo- ja proteiinimolekyylejä
kemian ja biokemian keinoin. Sen tärkein osa on perinnöllisyyteen
keskittynyt molekyyligenetiikka.
Biotekniikalla on sovelluksia lääketieteessä
(diagnostiikka, tautien hoito, lääkkeiden valmistus, geeniterapia),
maataloudessa (kasvin- ja eläinjalostus), elintarvike-, kemian- ja
puunjalostusteollisuudessa, kaivostoiminnassa, energian tuotannossa sekä jätteiden
käsittelyssä. Toistaiseksi tärkeimmät biotekniikan uudet sovellukset ovat
olleet lääketieteellisiä. Suurin hyöty on tiedon lisääntyminen sairauksien
perussyistä. Geenitekniikan avulla
voidaan kehittää tehokkaampia lääkkeitä ja tuottaa niitä esim. kasvi- tai
eläinsoluviljelmissä. Myös uusien, nykyistä tehokkaampien ja turvallisempien
rokotteiden kehittämisessä käytetään biotekn. menetelmiä. Biotekniikka on
parantanut mm. tartuntatautien ja perinnöllisten tautien sekä syövän
tunnistamista. Geeniterapialla tarkoitetaan ihmisen viallisen geenin korvaamista
uudella tai sen toiminnan estämistä.
Kasvibiotekniikan pisimmälle edennyt osa on mikrolisäys
eli kasvien suvuton lisääminen. Mikrolisäyksellä tarkoitetaan kasvisolujen
tai -solukkojen viljelemistä aseptisesti synteettisellä ravintoalustalla;
menetelmällä voidaan tuottaa yhdestä kasvista suvuttomasti sen identtisiä
kopioita. Mikrolisäyksen avulla tuotetaan mm. koristekasvien, vadelman, tyrnin,
koivun ja omenapuun taimia sekä siemenperunoita. Paljon tutkitaan myös
keinosiementen (somaattinen alkio) tuottamista. Uudenlaisia risteymiä voitaneen
tuottaa yhdistämällä eri kasvien protoplasteja (protoplasti). Geenejä
kasveihin voidaan siirtää esim. agrobakteerien avulla tai »ampumalla» niitä
soluihin. Biotekn. menetelmin pyritään mm. kehittämään myrkyille
vastustuskykyisiä ravintokasveja, kasveja, jotka pystyisivät hyödyntämään
ilmakehän typpeä, ja paremmin luonnon stressitekijöitä kestäviä
lajikkeita.
Kotieläinten jalostuksessa biotekniikan tärkeimmät
sovellukset ovat koeputkihedelmöitys, alkionsiirto, sukupuolen ennalta määrittäminen
ja perinnöllisen aineksen säilyttäminen pakastettuna. Alkionsiirrolla
tarkoitetaan hedelmöityneen munasolun jakamista identtisiksi soluiksi, joista
muodostuneet alkiot voidaan kasvattaa toisten eläinten kohduissa. Biotekn.
menetelmin pyritään mm. lihan ja maidon laadun parantamiseen, kasvunopeuden
lisäämiseen ja tautien vastustuskyvyn parantamiseen. Eläimillä on tehty myös
geeninsiirtotutkimuksia (transgeeninen eliö).
Elintarviketeollisuudessa biotekniikan avulla pyritään
tuottamaan tuotteita, jotka ovat ravintoarvoltaan ja muilta ominaisuuksiltaan
parempia kuin entiset. Tärkein menetelmä on elintarvikkeiden valmistuksessa käytettyjen
mikrobien perimän muuttaminen. Lisäksi pyritään uusien raaka-aineiden hyödyntämiseen
ja jätteiden hyväksikäyttöön. Puunjalostusteollisuudessa biotekniikkaa
voidaan käyttää esim. sellun valkaisussa, puun säilyvyyden parantamisessa ja
jätteen hyväksikäytössä ja kaivosteollisuudessa mineraalien irrottamiseen köyhistä
malmeista.
Biotekniikan nopea kehitys on aiheuttanut myös
keskustelua siihen liittyvistä eettisistä ongelmista ja riskeistä. Yksi suuri
riski on perimältään muutettujen eliöiden pääseminen luontoon. Yl. pidetään
biotekniikan tarjoamia mahdollisuuksia paljon suurempina kuin siihen
osa-alueita:
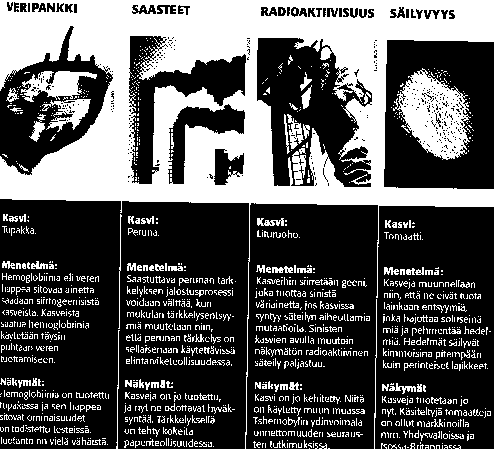
-tervetaimituotanto:
-lääkeaineitten tuotanto:
1.erytropoietiini (Huomen-lehmä), vaikuttaa
verenmuodostukseen ... ...normaalisti muodostuu munuaisissa...pyritään
tuottamaan maidossa.................
2.laktoferriini, tukee vastustuskyvyltään heikkojen
ihmisten puolustuskykyä (AIDS, syöpä- ym potilaat ?)............
3.kollageeni, korjaa kudosvaurioita, reumaa ym,
(Lapinlahti, Alapitkä ?)
(Laura Kulkas, MT 18. ja 22. 4. 1997)
.... eläinalkioon voidaan eristetty, haluttu geeni siirtää
mikroinjektiomenetelmällä: alkiosta otetaan kiinni, ja sen tumaan ruiskutetaan
geeniainesta
4.siirtogeenisiä kasveja voidaan käyttää rokotteiden
valmistuksessa (ja rokottamisessakin)... siirtämällä kasviin
taudinaiheuttajan se osa, joka käynnistäää elimistön puolustusreaktion,
voidaan rokote antaa syötävässä muodossa... kehitysmaihin ollaan kehittämässä
rokottamiseen tarkoitettuja siirtogeenistä perunaa ja banaania (MT 19.10 -96)
-tuotantoeläimet:
1.lohella muunnos, joka kasvaa nopeammin
2.kloonaus: esim Dolly-lammas: sai alkunsa utaresolusta ja
perimästään tyhjennetystä munasolusta, jotka fuusioitiin sähköllä (277
yritystä, 1 onnistui), tulos käyttäytyi kuin hedelmöittynyt munasolu. Tällä
menetelmällä voisi tuottaa perinnöllisesti tasalaatuisia karjoja!
(huom: urosta ei tarvittu)
3.Geenikartat--- markkerit, joista voidaan mikroskoopilla
tms havaita solusta (siemenneste
tms) perinnölliset ominaisuudet (geenit), mikä nopeuttaa jalostusta ja auttaa
risteytysvanhempien valinnassa (ilmiasusta ei yleensä voi vetää varmoja
johtopäätöksiä)
- kasvituotanto
-tuotantokasvit: Kaliforniassa tupakka tuottaa
malariatokotetta 250 kg/ha kuukaudessa !
Peruna
tuottaa albumiinia, jota tarvitaan veritulppapotilaiden hoitoon(Tieteen
Kuvalehti, liite 11/95)
EU:n alueella myönnetty (2/97) 759 lupaa geeniteknisiin
tutkimus- ja kehittämishankkeisiin . Eniten maissi, rypsi, rapsi, sok.juurikas, peruna
Usa: ssa (2/97) 23 lajiketta (kuusi maissia, viisi
tomaattia, neljä perunaa) , hankkeita melkein 2500... lajikkeista 17:llä
parannettu ominaisuus liittyy kasvinsuojeluun
1997 jo 15% USA:n soija-alasta GMO-soijaa (geneettisesti
manipuloitua organismia...)... muutenkin USA edistyksellisin
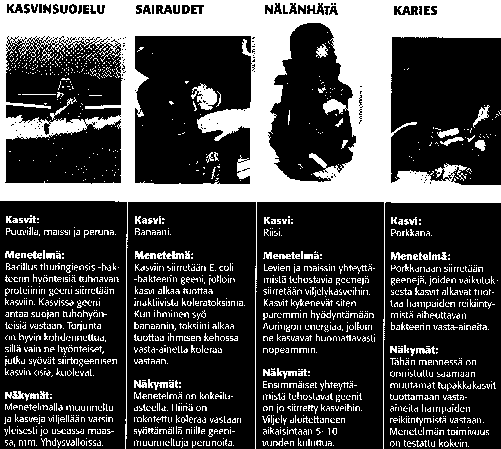
Bacillus thuringiensis-maabakteerin tuottamaa endotoksiinia on vuosikymmenet käytetty biologiseen perhostoukkien torjuntaan ruiskutteena. (hyväksytty yleisesti luomutuotannossakin) ... nyt maissiin siirretty ns. Bt-geeni, joka tuottaa maissikoisan tappavan luontoperäisen toksiinin ( Leipä Leveämmäksi 4/07)... onnistuu myös kaalikasveilla
useimmiten kasveilla tavoitellaan "resistenssiä ja
toleranssia" : vastustus- ja sietokykyä
-vastustusk.: lajike pystyy vastustamaan tuholaisia ja tauteja
-sietokyky useimmiten koskee torjunta-aineitten sietoa
esim soijan ja puuvillan viljelyssä paljon
kemikaaleja... GMO:lla pudotetaan puoleen ?
---- tuotanto taloudellisempaa ja ymp. ystävällisempää
joskus GMO:lla myös saavutetaan parempaa sadon laatua ja
varastokestävyyttä (esim tomaatilla)... myös esim "riisialueilla"
tms voisi ko. kasvia jalostaa ravitsemuksellisesti paremmaksi
(aminohappokoostumus yms).... (Tieteen Kuvalehti, liite 11/95)
-niinikään teollisuuden "palveluksessa" olevat
hiivat ym mikrobit ovat "räätälöidyt" tiettyyn tehtävään
sopiviksi (maidonjalostus,
alkoholiteollisuus...) esim EDAM-juuston juoksutuksessa käytetään kymosiinia,
jonka on tuottanut siirtogeeninen bakteeri
GEENINSIIRTO
ERÄS MAABAKTEERI, Agrobakterium tumefaciens aiheuttaa
aitosyöpää kasveissa (äkämiä, kasvaimia...)
siirtämällä DNA:staan kiihtyvää kasvua
aiheuttavan geenin kasviin
-geenisiirroissa käytetään agrobakteerikantaa, josta
tuo kasvua kiihdyttävä geeni
(bakteerin oma siirtogeeni) on korvattu kohdegeenillä ja tunnistegeenillä
(kyky tunkeutua isännän perimään on tallella) MT 19.10 -96
-agrobakteeriin halutut geenit saadaan esim. eshericia
coli-bakteerin viemänä (ks kuva)
-kolibakteerin plasmidiin (rengasrak DNA) joko
virusvektorin avulla tai suoraan
-DNA- avautumisessa ja sulkeutumisessa apuna
restriktioentsyymit, jotka kukin katkaisevat tai "liimaavat"
DNA-juosteen juuri tietystä kohdasta (MT 19.10. -96)... siirrettävän "hyöty"geenin
ohella liimataan samalla myös tunnistegeeni (esim antibiootin kestävyysgeeni),
jolloin siirtogeeniset kasvit voidaan erottaa antibioottia sisältävällä
kasvualustalla...
KUVA: BAKTEERIN PERIMÄN MUUTTUMISTAVAT
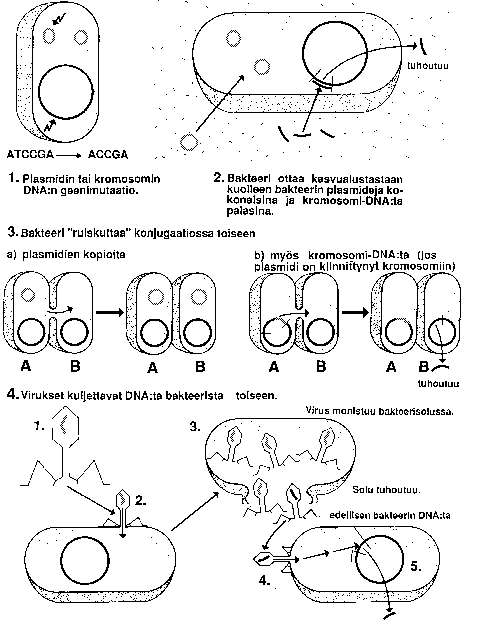
Kasveilla on geeninsiirtoja tehty:
yksisirkkaisilla: vehnä,ruis,ohra,riisi,maissi,hirssi
("partikkelipommituksella": geenipyssyllä ampuen,
kultahiukkasten pinnalla... , maissi jo 1990
kaksisirkkaisilla: lähinnä agrobakteerimenetelmällä
(kaksisirkkaisten luonnollinen loinen)
-hernekasveja: apila,herne,soija,sinimailanen
-koisokasvit: peruna,tomaatti,petunia,tupakka,munakoiso
-muita: rypsi,rapsi,kaali,porkkana,kurpitsa,salaatti, mansikka, viinirypäle, omena, mänty, kuusi, koivu, poppeli, päärynä
Luonnossa Agrobacterium tumefaciens -bakteeri infektoi
ainoastaan leveälehtisiä kasveja kuten tupakkaa, petuniaa, tomaattia ja rypsiä.
Se ei pysty yIeisimpiin viIjeIykasveihin vehnään, riisiin ja maissiin.
Vieraan DNA:n siirtämiseksi näihin kasveihin on käytettävä
aivan muita keinoja. Useimpia meneteImiä käytettäessä työ aIoitetaan
poistamalIa kasvin soluja suojaavat kerrokset. Tämä voidaan tehdä esimerkiksi
pilkkomalIa tehosekoittajaIla lehdet pieneksi siIpuksi. Sekoittajasta saatua
lehtipuuroa käsitellään tietyiIIä entsyymeillä, jotka hajottavat solujen
soluseinät. SoIuseinättömiä soIuja eli protoplasteja erottaa ulkomaailmasta
vain ohut solukelmu, mikä helpottaa DNA:n siirtoa soIuun. SoIukeImu voidaan
rikkoa moneIIa tavaIla. Yksi tapa On sekoittaa eri kasvien protoplasteja keskenään,
ja indusoida ne sulautumaan yhteen. Tällä menetelmäIIä on saatu aikaan muun
muassa pomaatti, tomaatti-peruna -hybndi, jossa on DNA:ta molemmista
emokasveista.
MeneteImällä perintöaineksen sekoittuminen on
sattumanvaraista, joten saatu pomaatti ei kyennyt lisääntymään. Yleensä
DNA-manipuIaatio on huomattavasti valikoivampaa ja se tähtää tiettyyn päämäärään.
Elektroporaatioksi kutsuttua menetelmää on käytetty menestyksekkäästi. Siinä
protoplastien ja puhdistetun DNA:n (joka on voitu ottaa mistä tahansa eliöstä)
seos pannaan sykkivään sähkökenttään. Sähkökenttä saa protopIastin
solukelmuun muodostumaan pieniä reikiä, joista DNA pääsee kulkeutumaan
soluun. Solukelmu voidaan rikkoa myös kemiallisilla menetelmillä. Molemmissa
tapauksissa DNA saadaan kulkeutumaan solun tumaan, jossa se sulautuu kasvin
kromosomeihin. DNA voidaan saada soluun myös ohuiIla injektioneuloilla, jotka
työnnetään varovasti solukelmun läpi. On olemassa myös hienostuneempia
menetelmiä, joilla DNA saadaan suoraan solun tumaan.Seuraava ongelma on saada
herkät protoplastit kehittymään kokonaisiksi kasveiksi. Useat protoplastit
vahingoittuvat fysikaalisissa, kemiallisissa ja sähköisissä käsitteIyissä.
Toiset protoplastit saadaan kasvamaan helposti, kun taas toiset eivät kehity
laisinkaan.Vaikeus piileekin kasvun alkamisessa, soIuseinän rakentamisessa
uudelleen - tutkijat eivät vielä kokonaan ymmärrä tätä
prosessia.Geeniteknikkojen ensimmäinen haaste oli saada aikaan
rikkakasvimyrkkyjä kestäviä kasvilajikkeita. Kasvimyrkyillä voidaan vähentää
rikkaruohoja, minkä ansiosta viljelykasvit saavat mahdollisimman paljon valoa
ja ravinteita. Glyfosaatti - kauppanimet Roundup ja Tumbleweed - on maanviljelijöiden
ja puutarhureiden suosima rikkaruohomyrkky. Se toimii estämällä erään
kaikissa kasveissa
olevan entsyymin toiminnan. Kasvi ei voi kasvaa ollenkaan
ilman tätä tärkeää entsyymiä. Glyfosaatti tappaa useimpien muiden
kasvimyrkkyjen tavoin valikoimatta kaikki kasvit niin rikkaruohot kuin
viljelykasvitkin. Siksi myrkky on levitettävä ennen kuin viljelykasvit
nousevat maasta. Tällöin myrkyn vaikutus ei ole niin tehokas kuin silloin, kun
sitä levitettäisiin myös kasvukauden aikana,jolloin rikkaruohotkin kasvavat
parhaiten.Myrkytyksen tehostamiseksi tutkijat ovat kehittäneet glyfosaattia
sietäviä kasvilajikkeita. Tähän tarvittava geeni saatiin glyfosaatille
resistentistä E. coli -suolistobakteerikannasta. Tutkijat eristivät geenin,
joka koodaa vastustuskykyyn johtavaa entsyymiä, ja siirsivät sen
Agrobacteriumin avulla tupakkaan, rypsiin ja petuniaan. Geneettisesti käsitellyt
kasvit kukoistivat rikkaruohomyrkytyksistä huolimatta. Kasvimyrkkyjen
sietokykyyn johtavat geenit voidaansiirtää myös perunaan ja tomaattiin.
Myrkynsietokyky mahdollistaa viljelykasvien kasvun ilman ravinnekilpailua
rikkaruohojen kanssa, mikä johtaa suurempiin satoihin pinta-alaa kohti. Ympäristönsuojelijat
ovat huolestuneita kasvimyrkkyjen kasvavasta käytöstä ja ympäristön
kemikalisoitumisesta.
- .
-
Geenitekniikka voi tehdä maatalouden käyttämät
erilaiset tuholaismyrkyt täysin tarpeettomiksi. Eräs Bacillus thuringiensis
-bakteerin (B.t) tuottama toksiini tappaa perhostoukkia, mutta ei vaikuta
lainkaan muihin hyönteisiin tai kasveihin. 1987belgialaiset ja yhdysvaltalaiset
tutkijat eristivät tästä bakteerista sen B.t.-toksiinia koodaavan geenin. Tämän
jälkeen geeni siirrettiin tupakantaimiin Agrobacterium tumefaciensin
Ti-plasmidin avulla. Kasvit alkoivat tuottaa toksiinia, joka tappoi niitä syövät
toukat. Myös virukset aiheuttavat merkittäviä satovahinkoja. Niitä vastaan
ei kuitenkaan ole olemassa minkäänlaisia kemiallisia - eikä muitakaan -
torjuntakeinoja. Ehkäpä geenitekniikka antaa keinot suojata viljelykasvit myös
virusten aiheuttamilta sairauksilta. Tupakan lehtiin läikkiä aiheuttava
tupakkamosaiikkivirus rakentuu proteiinikuoren sisään suljetusta RNA-kierteestä.
Jos pelkkä viruksen
proteiinikuorta
koodaava geeni siirretään tupakan soluihin, tuloksena saatu kasvi on immuuni
mosaiikkivirukselle. Sama tulos saatiin, kun tomaatintaimia käsiteltiin
vastaavalla tavalla. Tutkijat eivät vielä tarkkaan tiedä mistä tämä
johtuu. Geenitekniikalla voidaan myös parantaa kasvien ravintosisältöä.
Proteiinien
puute on tavallisin aliravitsemuksen muoto - etenkin kehitysmaissa, joissa usein
jopa 80 - 90 % väestön proteiinintarpeesta tyydytetään kasviravinnolla.
Ihmiselimistö kykenee itse tuottamaan vain kymmentä niistä 20 aminohaposta,
joita tarvitaan arvokkaiden proteiinien tuottamiseen. Loput aminohapot on
saatava ravinnon kautta. Useimpien viljelykasvien aminohapposisältö on varsin
alhainen. Kasvit varastoivat yleensä proteiinit siemeniinsä (papuihin, jyviin
ja pähkinöihin). Ravintokasvien lehtiä, varsia ja juuria ei usein käytetä
ollenkaan ravinnoksi. Kasvin ravintoarvoa voidaan parantaa saamalla ne
proteiinit, joissa on paljon ihmiselle välttämättömiä aminohappoja,
kulkeutumaan tehokkaasti kasvin siemeniin. Esimerkiksi monissa hernekasveissa,
(mm. pavuissa, herneissä ja linsseissä) joista pääasiallinen ravinto koostuu
suuressa osassa Etelä-Amerikkaa ja Afrikkaa, on erittäin vähän elimistölle
välttämättömiä, rikkiä sisältäviä aminohappoja. Tilanteen epätasapainoa
voidaan korjata liittämällä hernekasveihin parapähkinän siemenproteiineja
(joissa kyseisten aminohappojen osuus on jopa 25 % ) koodaavia geenejä. Toiseen
perusravintokasviin, perunaan, on siirretty keinotekoinen geeni, joka koodaa
useiden välttämättömien aminohappojen tehokasta tuotantoa. Geeni
valmistettiin geenikoneella, ja se liitettiin perunaan Agrobacteriumin
Ti-plasmidin avulla. On laskettu, että uusi geeni kaksinkertaistaa perunan
ravintoarvon. Monia kasveja viljellään niistä saatavien öljyjen ja rasvojen
takia.
Palmuista
saadaan raaka-aineita saippuan ja pesuaineiden valmistamiseen. Oliivi-,
auringonkukka-, maapähkinä ja soijaöljyjä käytetään elintarvikkeina.
Teollisuus käyttää rypsi- ja risiiniöljyjä voiteluaineina. Kaikki öljyt
ovat triglyseridejä. Ne rakentuvat kolmesta pitkästä rasvahappomolekyylistä,
jotka ovat kiinnittyneet toisesta
päästään
glyseroliin haarukan piikkien tavoin. Kasvin rasvahappojen tyyppi ja koostumus
riippuvat siitä, mitä entsyymejä kasvisolussa on. Geenitekniikka mahdollistaa
geenien siirtämisen kasvilajista toiseen, minkä ansiosta on mahdollista tehdä
kasveja, jotka voivat tuottaa mitä tahansa öljyä.
Jos tutkijat
esimerkiksi tietäisivät, miten kaakaopensas säätelee kaakaovoin tuotantoa,
he voisivat siirtää kyseiset geenit soijapapuun. Geenit koodaisivat
rasvatuotantoa ohjaavia entsyymejä. Soijapapua voidaan viljellä lauhkealla vyöhykkeellä,
ja siitä saatava kaakaovoi olisi huomattavasti »aitoa" kaakaovoita
halvempaa suklaan raaka-ainetta. Jos vastaavasti rypsi saataisiin tuottamaan
palmu- ja kookosrasvaa, trooppisten viljelykasvien tarve pienenisi olennaisesti.
.
Kasvisolut: uudistuminen, muuntelu ja viljely
Kotipuutarhurit kloonaavat jatkuvasti kasvejaan. Pensaasta
tai kukasta leikatusta oksasta tai versosta saadaan juurruttamisella kehittymään
uusi, kokonainen kasviyksilö, joka on lähes täysin samanlainen kuin emokasvi.
Tämä ei vaikuta ihmeelliseltä, mutta kyseessä on melkein sama ilmiö kuin
se, että amputoidusta jalasta voitaisiin kasvattaa täydellinen ihmisklooni.
Tutkijat kutsuvat kasveille ominaista kykyä kehittyä pienestä palasta
kokonaiseksi kasviksi totipotenssiksi. Bioteknologit ovat kehittäneet taitoa
kloonata kasveja vielä askeleen pidemmälle. He ovat jakaneet kasvin pienimpiin
mahdollisiin osiinsa, soluihin. Saamansa solut be ovat saaneet kasvamaan uusiksi
kasviyksilöiksi. Käytännön ongelma on saada uudistumisprosessi tapahtumaan
ehdottoman steriileissä oloissa. Hyvin pieni pala kasvisolukkoa (noin 0,5
kuutiosenttiä) puhdistetaan kemiallisesti, jolloin sen pinnalla olevat
mikro-organismit kuolevat.
Tämän jälkeen solukkoa käsitellään tietyillä
entsyymeillä, jotka hajottavat sen kiinteät rakenteet, esimerkiksi
selluloosan. Lopputuloksena saadaan liuos, jossa on tuhansia yksittäisiä
kasvisoluja. Solut voidaan saada kasvamaan steriilissä nesteessä lasipulloissa
tai petrimaljoissa, joihin lisätään solujen tarvitsemia ravinteita. Yhdessä
astiassa voidaan kasvattaa jopa satojatuhansia soluja, joista kukin kykenee
kehittymään kokonaiseksi kasviksi.
Kasveja uudistetaan kasvattamalla kasvisoluja hyytelömäisessä
aineessa, joka sisältää tiettyjä kasvuhormoneja ja ravinteita. Solut ovat lähes
täysin värittömiä, ja ne kasvavat aluksi muodottomana möykkynä, jota
kutsutaan kallukseksi. Parin päivän tai viikon kuluttua voidaan astioissa jo
havaita pikkuruisia vihreitä versoja, lehtiä ja juuria. Tätä prosessia
kutsutaan somaattiseksi embryogeneesiksi - alkionmuodostukseksi somaattisesta
solukosta. Alkioiden lukumäärä voidaan kymmenkertaistaa johtamalla solukon läpi
heikkoa sähkövirtaa 9 voltin paristosta, joka on kytketty kahden johtimen
kautta solukkoviljelmään. Solukon sterilisoimisen jälkeen
toinen paristosta lähtevä johdin kiinnitetään
kallukseen ja toinen hyytelömäiseen kasvualustaan. Tutkijat uskovat, että sähkövirta
antaa vielä muodottomalle kallukselle tunteen suunnasta. Kehittyneet
pienoiskasvit siirretään seuraavaksi omiin pieniin säiliöihinsä, joissa ne
jatkavat kehitystään. Muutaman viikon tai
kuukauden viljelmässä olon jälkeen kasvit ovat riittävän suuria ja
vahvoja istutettavaksi kompostiin. Siellä niiden annetaan kasvaa ja
vahvistua ennen kuin ne voidaan istuttaa tavalliseen
maahan. Kasvien kapselointi elatushyytelöä sisältävään suojakuoreen jo
alkiovaiheessa on mielenkiintoinen vaihtoehto asteittaiselle kasvatukselle.
Keinotekoiset kapselisiemenet voidaan istuttaa suoraan maaperään
erikoiskoneiden avulla. Yksittäisten solujen kloonaaminen on erittäin nopea
tapa lisätä taimien lukumäärää. Nykyään uutta kasvilajia kehitettäessä
sitä voidaan toimittaa suuria määriä viljelijöiden käyttöön
huomattavasti nopeammin kuin ennen.
5. kenttäkoetekniikka
6. matemaattinen analyysi
F-testi ja T-testi
varianssi s2 = S( (xi-xka )2 ) / (n-1) , jossa xka = keskiarvo
1.
havainnot:
keskiarvo
varianssi
a
19
14
15
17
20
17
6,5
b
23
19
19
21
18
20
4,0
yhteinen keskiarvo ja varianssi.........
18,5
5,25
2.Oletetaan hypoteesi: ovat samasta populaatiosta... eli
on yhteinen varianssi ja keskiarvo olemassa:
-tämä yhteinen keskiarvo = 18,5
-tämä yhteinen varianssi = 5,25
3. Keskiarvojen vaihtelua kuvaava keskiarvojen varianssi
tulee kaavasta:
s2x = ((xb-x)2 + (xa-x)2) / (2-1) =((17-18,5)2 +
(20-18,5)2) /
1 = 4,5
(keskiarvon keskihajonta = neliöjuuri 4,5 = 2,1)
Jos tämä arvo on kovin suuri, se osoittaa käsittelyjen
(lajikkeet tms.) poikkeavan toisistaan paljon
Jos lisäksi aiemmin lasketut A:n ja B:n varianssit ovat
pienet (ja yhteinen var. pieni... 5,25) niin tämä johtaa päätelmään, että
lajikkeilla on todellista eroa
4. Estimoidaan s22, eräänlainen kokonaisvarianssi = n*s2x =
5*4,5 = 22,5
F=s22/ s21
eli 22,5 / 5,25, siis lajikkeiden väl. eroista aiheutuva / laj.
sisäisestä hajonnasta aiheutuva tekijä
MITÄ SUUREMPI F-ARVO, SITÄ VARMEMMIN ON TODELLISIA EROJA KÄSITTELYJEN VÄLILLÄ
PARIVERTAILUSSA
t = (xa-xb) /
sd,
missä sd =
"keskiarvojen erotuksen keskivirhe" =
V ((s2a + s2b) / n) =
V ((6,5 + 4,0) / n) =
1,449
( V=neliöjuuri)
t = | 17-20 | / 1,449 =
2,07
HUOM: jakaja sitä suurempi, mitä suurempi on kokeen ympäristö- ja
satunnaistekijöistä johtuva epävarmuus, siis hajonta, jota kaavassa kuvaavat
A:n ja B:n varianssit. Tällöin t:n arvo tulee pieneksi. Taas, mitä suurempi
on osoittaja, eli käsittelyjen (lajikkeitten tms) välinen ero.... sitä
suurempi t:n arvo, mikä osoittaa kokeen tuloksen merkitsevyyttä