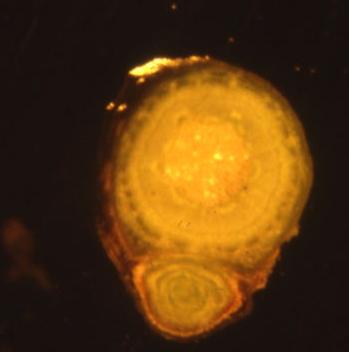
Juuren tehtävät
-
kiinnittää,tukee,ottaa vettä ja ravinteita, hengittääkin

KASVIBIOLOGIA kasvin rakenne ja elintoiminnat, Matti Pekkarinen (kirjallisuus: Pohjola-Valanne: kasviopin ja ympäristönsuojelun perusteet ammatillisille oppilaitoksille)
1. Ulkorakenne (organologiaa)
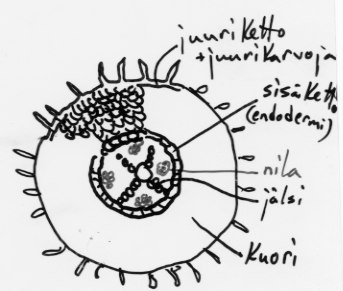
JUURI
|
Juuren tehtävät
|
|
|
|
VARSI (oikealla poikkileikkaus koiranheisi- puusta) |
|
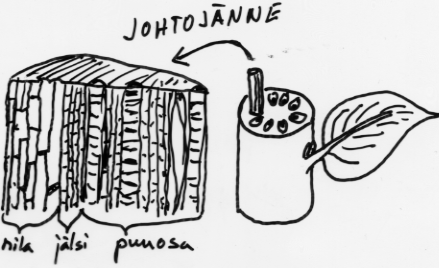
Varren tehtävät: -kannattelee lehtiä ja kukkia, johtaa aineita (nesteitä) alas ja ylös Varren pituuskasvu riippuvainen valonsaannista (hormonaalinen säätely)
|
 |
LEHTI
Lehden tehtävät
yhteyttäminen (ml valon "pyydystäminen")
haihduttaminen (välttämätön ravinteidenotolle maasta: nestevirtaus läpi kasvin)
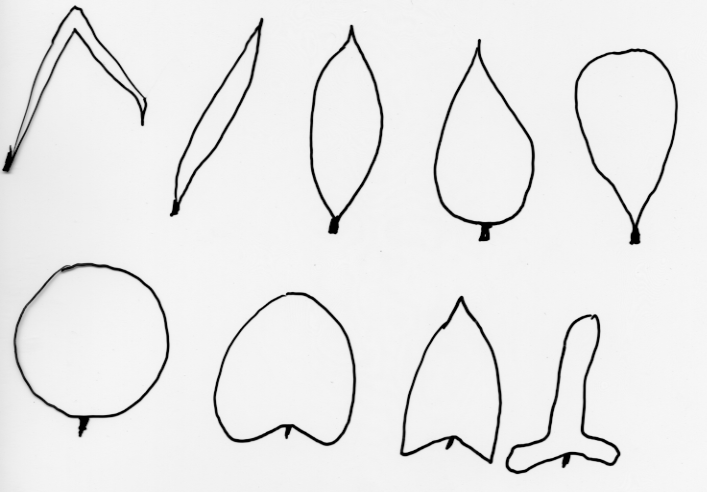
| Lehtimuotoja: tasasoukka, suikea, soikea, puikea, vastapuikea, pyöreä, munuaismainen, nuolimainen, keihäsmäinen |
 |
|
KUKKA |
 |
Myös kukka on lehdistä muodostunut:
kukkalehdet:
kehälehdet (=verho-ja terälehdet) ja
siitoslehdet (=hede-ja emilehdet)
heteen osat: palho,ponsi
emin osat: sikiäin,vartalo,luotti
Kukan tehtävät:
tuottaa siementä, jatkaa sukua
hedelmöityksen jälkeen muuttuu emin sikiäin hedelmäksi ja sen sisällä olevat siemenaiheet siemeniksi
|
Kukan rakenne
|
kukinto-ja kukkatyyppejä runsaasti: pääasiallisena perusteena kasvien taksonomiassa,luokittelussa sukuihin ja lajeihin
|
Kukinto... joillakin lajeilla (pienikukkaisia) kukat ovat ryhmittyneet enemmän tai vähemmän tiiviiksi ryhmiksi (terttu,tähkä...mykerö...sarja...kerrannaistähkä...röyhy)
2. KASVIN KEHITYSVAIHEET
ITÄMINEN
Itämisen edellytykset:
itämiskypsyys- kaikki siemenet eivät ole heti itämiskelpoisia (keväällä itävät rikkalajit, myös kevätviljoilla) - ITÄMISLEPO ELI DORMANSSI. Kylmäkäsittely (stratifikaatio) murtaa itämislevon
siemenen ikä. viljoilla 2-3 vuotta itävyys hyvä (samoin luonnonkasveillakin?). Jopa 50v voi osa siemenistä säilyä itämiskykyisinä. Riippuu lajista ja oloista.
ulkoiset edellytykset
vesi, kosteus
lämpötila
happi: tallaaminen ja sateisuus vähentävät
valo: siemen aistii, missä syvyydessä se on. Pienet siemenet vaativat valoa, isot eivät.
vanhoilla vaatimukset pienemmät
Itämisessä siemenen kuori kehittää amylaasi-entsyymiä, joka kulkeutuu vararavintosolukkoon. Siellä se muuttaa tärkkelystä sokereiksi, jotka kulkeutuvat alkioon. Alkio käyttää sokereita kasvun energian lähteenä.
VILJA JYVÄ:
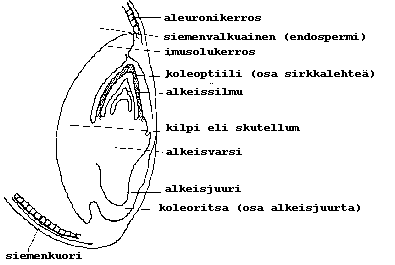
|
TAIMIVAIHE JA VEGETATIIVINEN KASVU |
 |
KUKKIMINEN
suvullista lisääntymistä
tietty vegetatiivinen vaihe läpikäytävä ensin
pituus vaihtelee----eräillä yksiv rikkakasveilla todella lyhyt
päivänpituus tärkeä säätelijä.......Lyhyenpäivänkasvit ja pitkänpäivänkasvit
helle hoputtaa.......kukkiminen tietyn lämpösumman täytyttyä
monet lauhkean ja viileän vyöhykkeen kasvit vaativat vernalisaation
HEDELMÖITYS + SIEMENENMUODOSTUS
hedelmöittymistapa vaihtelee: itsesiittoisia ja ristisiittoisia
myös apomiktinen lisääntyminen
hedelmöittyminen kevätviljoilla tapahtuu tähkälletulon aikoihin
siementen määrä (tähkylöiden määrä) riippuu paljon ympäristötekijöistä
runsas typensaanti näyttää lisäävän jyvälukua
viileässä enemmän tähkylöitä (ja siten myös jyviä)
kuivuus vähentää selvästi jyvälukua ---lyhyet tähkät
pitkä päivä hoputtaa--laskee hieman jyvälukua pohjoisessa
jyvän muodostus-eli täyttymisvaihe kestää kukkimisesta tuleentumiseen saakka
alkuosa kasvun aikaa:jyvään varastoituu tärkkelystä ja muita aineosia
kasvualustan kosteus ja typen riittävyys määräävät tämän vaiheen pituutta ja sadon suuruutta
loppuosa on passiivisen tuleentumisen aikaa : kuiva-ainesato ei enää kasva...jyvän kuiva-ainepitoisuuden nousu johtuu veden haihtumisesta joka taas on riippuvainen ympäristöoloista: lämmöstä ja ilmankosteudesta
KASVUAIKA KYLVÖSTÄ TULEENTUMISEEN voi samallakin lajikkeella vaihdella melkoisesti. Lämpösumma on jo parempi kasvukauden mittari kuin päivien lukumäärä. Muiden tekijöiden (typen-ja vedensaanti ja päivänpituus) vaikutus sekoittaa asiaa siten ,ettei lämpösummakaan ole aivan yksiselitteinen. Lajien ja lajikkeiden välisien kasvuaikavaatimuserojen ilmaisijana lämpösumma on hyvä mittari.
Viljelyvarmuustekijänä kasvuaika olisi saatava lyhyeksi. Kasvuaikaa lyhentävät tekijät vaikuttavat kuitenkin usein satoa alentavasti (jäähän osa kasvukautta hyödyntämättä) Siksi käytännössä pitkä kasvuaika - olipa se sitten lajikkeesta tai sää- ym. tekijöistä riippuvainen - johtaa pääsääntöisesti laarien täyttymiseen, lyhyt niiden jäämiseen vajaiksi.
VILJAN KEHITYSASTEET, useita jaottelutapoja...
1.ITÄMINEN
2.ORASVAIHE
orastumisesta korrenkasvun alkuun , samalla tähkänmuodostuksen alkuun
pensastuminen
3.KORSIVAIHE
jyvien lukumäärä määräytyy
pensastuminen loppuu (ellei harvaa tai lakoa)
korsien pituuskasvua
kasvimassan lisäys nopeaa
4.JYVÄNMUODOSTUS
tähkälletulon aikoihin tapahtuu kasvin kukissa hedelmöittyminen ja jyvän kasvu alkaa
jyvään varastoituu tärkkelystä
jyvän kosteus vähenee,alussa kasvi poistaa vettä aktiivisesti=aktiivinen tuleentuminen --- FYSIOLOGINEN TULEENTUMISVAIHE,jolloin kosteutta noin 40%
tämän jälkeen passiivisen tuleentumisen vaihe,jolloin jyvän kosteuden muutokset ovat riippuvaisia sääoloista
yleinen jaottelu eri tuleentumisasteisiin on seuraava:
MAITOTULEENTUMISASTE---KELTATUL.ASTE---TÄYSTUL.ASTE---YLITUL.ASTE
tuntomerkkeinä halmeen yleisväri, jyvän kiinteys, kovuus, jyvän väri, lehtien ja korren väri
keltatul. aste ja em. fysiologinen tuleentumisaste ovat jokseenkin samaan aikaan. Tällöin on jyvän kuiva-ainemäärä jo täysin muodostunut, eikä "sirppihallaa" synny.
| TULEENTUMISVAIHEIDEN TUNTOMERKKEJÄ: |
 |
MAITOTULEENTUNUT: alalehdet keltaisia, ylälh vihreitä ,jyvä maitomainen, latvapäästä kellertyvä, solmut ja korret vihreät, tuoreet, kosteus yli 50%
FYSIOLOGINEN TULEENTUNEISUUS: kosteus 35-40 % , jyvä vetinen, jonkin verran taikinamainen tai jauhomainen, vihreä jyvistä häviämässä, jyvä taipuva (vehnä, ruis: halkeaa) korret ja solmut kellastuvat
KELTATUL: kosteus noin 30%,jyvä sitkeä, jauhomainen ja taikinamainen, taittuu poikki, murtopinta epäsäännöllinen, halme kokonaan keltainen, alasolmut kutistuvat
TÄYSTUL: kosteus 20-15%,jyvä lasimainen/jauhomainen, kova, tähkät nuokkuvat , halme harmaantuu, solmut kutistuvat
YLITUL: kasvusto "ränsistynyttä", harmaata, korsi hauras --- korret taittavat, tähkiä putoilee, idäntäriski suuri
lisääntyminen
a) Suvuton lisääntyminen
ei hedelmöitystä -> ei muuntelua -> perimä säilyy samana, jälkeläiset emonsa klooneja
a) kasvullinen lisääntyminen: peruna, juolavehnä, marjapensaat. Esim perunalajike on klooni: yhdestä emoyksilöstä "monistettu"
b) lisääntyminen siemenestä, joka syntyy ilman hedelmöitystä (apomiktinen lisääntyminen, esim nurmikkalajeilla)
b) Suvullinen lisääntyminen:
hedelmöityksen tulos: kahden solun perinnöllinen aines yhdistyy
itsesiitos tai ristisiitos
Itsesiitos
itsesiitos: varma pölytys, ei uutta geeniyhdistelmää (ohra, kaura, vehnä, herne)
Ristisiitos
ristisiitos tuottaa uusia yhdistelmiä (ruis, apila)
ristisiitoksessa useita pölytystapoja: yleisimmät tuuli- ja hyönteispölytys
ristisiittoisilla mekanismeja, jotka estävät itsesiitoksen :
kukkien yksineuvoisuus: "poika- ja tyttökukat erikseen" (maissi, havupuut) ... tällöin suurella todennäköisyydellä hedelmöittävä siitepöly tulee eri yksilöstä (kaksineuvoisessa kukassa on hede- ja emilehdet samassa kukassa)
kasvien kaksikotisuus: on erikseen "tyttö- ja poikakasveja"! Mm tyrni, pajut, haapa. Niillä tapahtuu aina 100% varmuudella eri yksilöiden välinen ristisiitos (yksikotiseksi sanotaan kasvityyppiä, jolla samassa yksilössä on heteet ja emit)
Jos kasvi on yksikotinen, ja kukat kaksineuvoisia, on itsesiitoksen estoon muita keinoja: heteiden ja emin eriaikainen kehitys, kemialliset (=geneettiset) estomekanismit) ja (varsinkin tuulipölytteisillä) jo itsesiitoksen suuri epätodennäköisyys. Esim ruispellossa kun lentelee miljoonien kukkien siitepölyä yhtä aikaa.
Hedelmöitys
siitepöyhiukkasen tuma (oik. 2 kpl) kulkeutuu emin siemenaiheen alkiorakkoon. Toinen yhtyy munasolun tumaan (n+n=2n), josta syntyy uuden siemenen alkio. Toinen yhtyy alkiorakon keskustumaan (2n+n=3n), josta syntyy uuden siemenen vararavintosolukko.
3. Yhteyttäminen
yhteyttäminen, fotosynteesi, tapahtumasarja, jossa vihreät kasvit (siemenkasvit, sanikkaiset, sammalet, levät) ja eräät bakteerit sitovat Auringon valoenergiaa kem. energiaksi. Yhteyttämisen lähtöaineina ovat vesi ja ilmakehän hiilidioksidi. Reaktiotuotteena syntyy orgaanista, runsaasti energiaa sisältävää sokeria sekä happea:
6 CO2 + 6 H2O + valon energiaa ------- C6H12O6 + 6 O2 eli hiilidioksidi + vesi ---- glukoosi + happi. Yhteyttäminen on elämän perusta maapallolla. Vihreiden yhteyttävien kasvien sitoma energia välittyy ravintoketjussa toisenvaraisille (heterotrofisille) eliöille. Ilmakehän happi on peräisin yhteyttämisestä, ja vihreät kasvit pitävät yllä ilmakehän happi–hiilidioksiditasapainoa. Merien planktonlevät tuottavat hapesta huomattavan osan. Kivihiili, öljy ja maakaasu ovat biol. materiaalin hajoamistuotteita, jotka ovat peräisin yhteyttävien eliöiden vuosimiljoonia sitten synnyttämistä yhdisteistä.
Yhteyttäminen tapahtuu kasvisolujen viherhiukkasissa (kloroplasteissa), joita on runsaasti lehdissä. Siinä voidaan erottaa kaksi vaihetta: valoreaktiot, joiden edellytyksenä on valoenergian saanti, sekä pimeäreaktiot (Calvinin kierto), jotka seuraavat välittömästi valoreaktioita, mutta ovat mahdollisia myös pimeässä.
Valon vaikutuksesta lehtivihreä (klorofylli) virittyy, mikä mahdollistaa elektronien irtoamisen ja siirtymisen toiselle molekyylille. Yhteyttämisen ns. elektroninsiirtoketjussa elektronin vastaanottava molekyyli pelkistyy ja luovuttava molekyyli hapettuu. Hapetus–pelkistysreaktioissa vapautuva energia varastoituu ATP:hen (adenosiinitrifosfaattiin), jota syntyy ADP:stä (adenosiinidifosfaatista) ja epäorgaanisesta fosfaatista ja joka on runsasenerginen yhdiste. Siitä vapautuu kahden uloimman fosfaattiryhmän irtautuessa paljon energiaa, ja sitä käytetään solussa energian siirrossa ja tilapäisvarastona. Lehtivihreämolekyyli saa puuttuvat elektronit vesimolekyylistä, josta valon vaikutuksesta vapautuu happea, vetyioneja H+ ja elektroneja (e-). Viimeisenä elektronien vastaanottajana toimii NADP (nikotiinihappoamidiadeniinidinukleotidifosfaatti), joka pelkistyy NADPH2:ksi ja joka solussa osallistuu yleisesti hapetus–pelkistysreaktioihin. Pelkistyessä siihen liittyy vetyä ja elektroneja, ja hapettuessa se luovuttaa niitä. Valoreaktioissa on siis kolme rinnakkaistapahtumaa: ATP:n muodostuminen (fotofosforylaatio), veden hajoaminen (fotolyysi) ja vedynsiirtäjän, NADP:n, pelkistyminen.
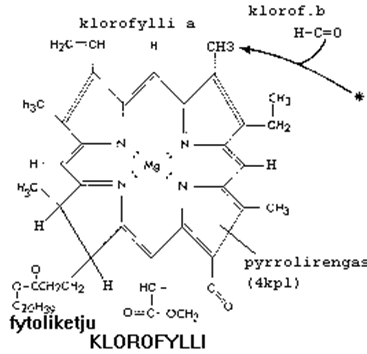
Valoreaktioiden perustuotteet NADPH2 ja ATP käytetään pimeäreaktioissa hiilidioksidin sitomiseen ja pelkistämiseen. Hiilidioksidin, jota versokasvit ottavat ilmarakojen kautta, vastaanottajana on 5-hiilinen sokeri, ribuloosi-bifosfaatti (RuBP). Tätä reaktiota katalysoiva entsyymi on luonnon yleisin valkuaisaine. Ensimmäinen havaittava välituote on 3-hiilinen sokeri, fosfoglyserolihappo, joka useiden välivaiheiden kautta muuttuu glukoosiksi. Reaktiotuotteista muodostuu solun muita tärkeitä yhdisteitä: pidempiketjuisia hiilihydraatteja, rasvahappoja ja aminohappoja. Hajoavan ATP:n energia varastoidaan pysyvämpiin energian varastomolekyyleihin kuten tärkkelykseen tai rasvoihin. Yhteyttämisessä sokeriin sitoutunutta energiaa vapautuu hengityksessä ja käymisessä.
Kemosynteesissä eräät bakteerit (esim. rikki- ja nitrifikaatiobakteerit) sitovat hiilidioksidia ilman Auringon valoenergiaa. Hiilidioksidin pelkistämiseen tarvitsemansa energian ne saavat hapettamalla yksinkertaisia epäorgaanisia yhdisteitä, kuten ammoniumsuoloja tai sulfideja.
Epäorgaanisen typen ja rikin muuttamista orgaanisiksi yhdisteiksi sanotaan joskus myös yhteyttämiseksi (assimilaatioksi). Typensidonnassa käytetään lähtöaineena kaasumaista typpeä (N2).
4. Kasvutekijät ks kappaleet 1 ja 2 täältä
5. Kasvin vesitalous ja ravinteiden otto
Kasvin vesipitoisuus ja veden tarve
vettä yl. noin 70-80 % elävissä ja kasvavissa kasvinosissa
monet hedelmät sisältävät yli 90 % vettä
siemenet ja itiöt vain noin 10 %
vesi on tarpeen aineenvaihdunnassa,j oka tapahtuu aina vesiliuoksissa.. aineiden kuljetuksessa - vesi on välttämätön
kasvin nestejännityksen ylläpitäjänä - - - - "kasvin ryhti "
yhteyttämisen raaka-aineena myös pienehkö määrä vettä
Veden otto ja kuljetus
vedenotto perustuu diffuusioon ja osmoosiin
a) Diffuusio on itsestään tapahtuvaa aineiden sekoittumista , perustana molekyylien lämpöliike
tasoittaa vallitsevia väkevyyseroja.........nesteissä hidasta
b) Osmoosi on diffuusiota puoliläpäisevän (semipermeaabelin) kalvon lävitse
kasveilla kyse veden tunkeutumisesta puoliläpäisevän kalvon läpi laimeasta liuoksesta väkevään , ts. osmoosi pyrkii tasoittamaan väkevyyseroja. Solukelmu päästää lävitseen helposti vettä,mutta ei siihen liuenneita aineita
yleensä kasvin solunesteen suolaväkevyys on suurempi kuin juuria ympäröivän maanesteen.Tähän perustuu veden kulkeutuminen kasvin juureen.
osmoosiin perustuva vedenotto ei vaadi kasvilta energiaa
ravinnesuoloja kasvi ei saa osmoosin avulla , vaan niiden otto on juurisolujen työtä ja vaatii energiaa
Veden kulku kasvissa
juurikarvat ottavat tehokkaasti vettä (moninkertaistavat juuren pinta-alan)
vesi kulkeutuu osmoottisen imun ansiosta solu solulta juuren johtojänteisiin
johtojänteissä vesi nousee haihtumisimun (ja veden koheesiovoimien) ansiosta ylös lehtiin
kapillaari-ilmiö edesauttaa nousemista
vesi kohoaa myös juuriston solujen aktiivisen paineen,juuripaineen avulla
Veden haihtuminen
haihtuminen aikaansaa nestevirtauksen ja ravinteiden nousun verson ylimpiin osiin asti
haihtuminen sitoo myös lämpöä (vähentää haitallista kuumenemista hellekausina)
suurin osa haihtumisesta tapahtuu ilmarakojen kautta- - - - huulisolut säätelevät
liiallista haihtumista vastaan kasveilla on useita keinoja: ilmarakojen lukumäärä ja sijainti,lehtien koko ja muoto , vaha- tai karvapeite ,lehtien kokoonkäpertyminen jne
talvehtivia kasveja voi vaivata kuivuus varhaiskeväällä- - - - jääpolte
Kasvien ravinteiden otto
Kasvien ravinteet ovat maanesteessä pääasiassa ionimuodossa :
kationeina 85 % ravinteista: NH4 , K , Ca , Mg , Na , ( Cu , Mn , Zn , Fe ....)
anioneina 15 % : PO4 , BO4 , NO3 , SO4 ....
maahiukkasilla yleensä negatiivinen varaus ----- pystyvät sitomaan kationeja
KVK riippuvainen Ph:sta ------- happamassa pieni
kationien sidosvoimat erisuuruisia ; järjestys : Na , K , NH4 , Mg , Ca , H
happamassa H syrjäyttää muita , "varaa paikat"
anionit liuenneina maanesteeseen :NO3 helppoliuk , fosfaatit vaikealiuk
"vaihtuvana" posit.var.pinnoille: happamassa saviminer. ja orgaaninen aines
( ja Fe- ja Al-hydroksidit)
"anioninvaihtokapasiteetti" liukuva käsite: anionit muod. usein yhdisteitä maaaineksen kanssa- - - - liukoisuus vaihtelee
AINEIDEN KULJETUS
Kasveissa vesi ja siihen liuenneet ionit kulkevat johtojänteiden puuosassa. Orgaaniset yhdisteet (yhteyttämistuotteet) taas kulkevat nilaosassa.
Nilassa virtaavaa orgaanisten yhdisteiden vesiliuosta sanotaan nilanesteeksi. Se sisältää kuiva-ainetta 10-30 %.Siitä valtaosa on sakkaroosia.
Lisäksi siinä on aminohappoja ja fosfaatteja,kaliumia ja kasvuhormoneja..
Nesteen virtausnopeudeksi on mitattu pajuilla 30-100 cm/h.
Virtailu tapahtuu aina tuottavalta solukolta kuluttavalle päin. Esim.keväällä puilla tapahtuu virtausta juurista silmuihin päin,kesällä päinvastaiseen suuntaan.
(nilanestetutkimuksissa käytetään "lehtitäitekniikkaa")
6. KASVIEN KASVUA SÄÄTELEVÄT AINEET (hormonit ja niiden inhibiittorit)
Tärkeimmät hormoniryhmät : auksiinit , gibberelliinit ja sytokiniinit
AUKSIINIT (kreikk. auksein: "kasvaa")
edistävät kasvisolujen pituuskasvua(lisäämällä soluseinien joustavuutta)
vaikuttavat myös solujen jakautumiseen,hedelmien kehittymiseen ja kypsymiseen
myös yhteisvaikutuksia muiden horm.kanssa .. mm.johtosolukon kasvun säätely,
varren haaroittuminen (apikaalisen dominanssin säätely)
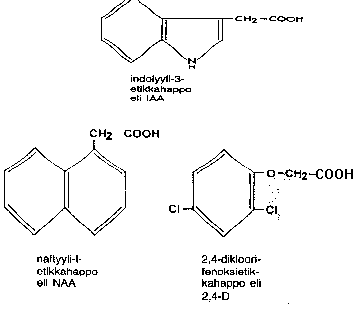
indolyylietikkahappo IAA ja fenyylietikkahappo PAA ym - yhteensä yli 10 yhdistettä
synteettisiä auksiineja,mm 2,4-dikloorifenoksietikkahappo eli 2,4-D ja 2,4,5-T rikkakasvintorjunnassa
kasvissa auksiinit syntyvät verson kärjessä,josta kulkeutuvat nilavirtauksessa alas
säätelevät juuristossa syntyvää sytokiniiniä -------- yhdessä luovat tasapainoisen kasvun kulun
GIBBERELLIINIT
YLI 50 ERI YHDISTETTÄ , yleismerkintä GAx , jossa alaindeksinumero = löytöjärjestys
yleisin GA3 eli gibberelliinihappo
gibberelliinit lisäävät solunjakautumista ja myös solujen pituutta
gibberelliinillä on valo- ja kylmäkäsittelyä korvaava vaikutus (itämislevon poisto siemenestä , samoin vernalisaation korvaajana syysyksivuotisilla)
vaikuttaa myös kukkimiseen , riippuu kasvilajista
voidaan myös säädellä hedelmien kypsymistä ja edistää partenokarppisten hedelmien syntyä (viinirypäleet ....)
oluen valmistuksessa GA3:lla parannetaan amylaasientsyymin tehoa idätysvaiheessa
SYTOKINIINIT JA ETYLEENI
sytokiniinit SUBSTITUOITUJA ADENIINEJA
vaikutus kohdistuu mitoosiaktiviteettiin so.lisäävät solujakoja
myös solujen laajuuskasvu lisääntyy ja vanheneminen hidastuu
etyleeniä syntyy kasvin vanhenevissa osissa,mm kellastuvissa lehdissä ja kypsyvissä hedelmissä ----"kypsymishormoni"
voidaan käyttää kaupallisesti esim vihreiden banaanien kypsyttämiseen
KLORMEKVATTI on käytännön kannalta merkityksellisin kasvunsääde
keksittiin 1950-luvun lopulla
toimii gibberelliinisynteesin estäjänä
ABSKISSIHAPPO estää lähes kaikkien kasvinosien kasvua, siementen itämistä ja silmujen puhkeamista, ts se edistää lepotilan eli dormanssin säilymistä
KUMARIINI on laktoniyhdiste,joka estää mm siementen itämistä,auksiinisynteesiä ja SH-entsyymien toimintaa
7. Perinnöllisyys (herediteetti) (ks myös täältä)
Toisilleen sukua olevat eliöyksilöt muistuttavat toisiaan enemmän kuin saman lajin yksilöt keskimäärin. Tämä kuuluu elämän perustavimpiin tunnusmerkkeihin. Perimää välittää sukupolvesta toiseen deoksiribonukleiinihappo eli DNA, joillakin viruksilla ribonukleiinihappo eli RNA. Bakteereilla DNA sijaitsee rengasmaisena molekyylinä solulimassa, muilla eli ns.aitotumaisilla eliöillä se on tuman kromosomeissa. Ennen solun jakautumista sen DNA-molekyylit kahdentuvat niin, että syntyvät tytärsolut saavat samanlaisen DNA-koostumuksen; Perinnöllisyys ja perimän siirtyminen uusiin soluihin ja sukupolvesta toiseen perustuu juuri nukleiinihapon kahdentumiskykyyn. Jossakin määrin perinnöllisyyttä välittävät myös kromosomista irrallaan olevat plasmidit sekä solulimassa olevat DNA:ta sisältävät soluelimet, kuten mitokondriot ja plastidit.
Perinnöllisyyden yksikkö on perintötekijä eli geeni. Saman geenin eri muotoja, sanotaan alleeleiksi (esim ihmisen silmän väriin vaikuttavasta geenistä). Jos kaksi kromosomistoa käsittävän eli diploidisen solun tai yksilön jonkin kromosomin jossakin geenikohdassa eli lokuksessa on sama alleeli kuin sen vastinkromosomin vastaavassa lokuksessa (so. samaa alleelia on solussa kaksi kappaletta), solu tai yksilö on kyseisen geenin suhteen samaperintäinen eli homotsygoottinen. Jos taas on eri alleelit, solu tai yksilö on kyseisen geenin suhteen eriperintäinen eli heterotsygoottinen. Täydellistä homotsygotiaa (kaikkien geenien suhteen) esiintyy monisoluisilla eliöillä hyvin harvoin jos koskaan. Jatkuvalla sukusiitoksella (esim itsesiittoisilla kasveilla) voidaan päästä hyvin lähelle sitä.
nukleiinihapot, DNA
Suurimolekyylisiä polynukleotideja, joihin sisältyy eliöiden perimä ja jotka ohjailevat valkuaisaineiden rakentumista soluissa. Ne muodostuvat yhdistymällä mononukleotideista. Nämä käsittävät yhden fosforihappotähteen, yhden pentoosisokerin (deoksiriboosi tai riboosi) ja yhden typpipitoisen puriini- tai pyrimidiiniemäksen. Puriineja ovat adeniini (A) ja guaniini (G), pyrimidiinejä tymiini (T), urasiili (U) ja sytosiini (C). Deoksiribonukleiinihapon eli DNA:n sokeriosat ovat aina deoksiriboosia, ja sen emäksiä ovat A, T, G ja C. Perättäisten emäsosien tietty järjestys ja sen säilyminen kunkin eliön DNA:ssa muodostaa perinnöllisyyden perustan. Jokainen kolmen emäksen sarja (tripletti eli kolmikko) merkitsee geneettisessä koodissa tiettyä aminohappoa. Muutaman sadan tripletin pituinen DNA-jakso merkitsee aminohapoista rakentuvaa valkuaisainetta.
nukleiinihapot, RNA
Valkuaisaineen valmistusta varten DNA:n kaksoisjuoste aukeaa jonkin geenin kohdalta, ja jommankumman juosteen rinnalle rakentuu sille komplementtinen ribonukleiinihappojuoste (lähetti-RNA). Ribonukleiinihapon (RNA) molekyylit ovat aina yksijuosteisia; niissä on sokeriosana riboosi ja emäksinä A, U, G ja C. Muut valkuaisaineiden synteeseihin osallistuvia RNA-tyyppejä ovat ribosomi-RNA:t ja siirtäjä-RNA:t.
tumanjakautuminen
Tumanjakautuminen on solunjakautumisen aloittava tapahtumaketju, jota on kaksi tyyppiä:
a) mitoosi, jossa syntyneet tytärsolut sisältävät täsmälleen samat kromosomit kuin emosolukin, ja meioosi, jossa tytärtumien kromosomien lukumäärä vähenee puoleen. Mitoosin esivaiheessa, profaasissa, pitkittäin kahdentuneet kromosomit alkavat tulla näkyviksi (valomikroskoopilla katsottaessa) kiertymisen aiheuttaman paksuuntumisen ansiosta. tumajyvänen ja -kelmu häviävät. Samanaikaisesti on sentrosomi jakautunut kahtia, ja sen puoliskot siirtyvät tuman vastakkaisille puolille. Tumasukkula alkaa muodostua niiden väliin. Keskivaiheessa, metafaasissa, yhä selvemmin näkyvät kromosomit siirtyvät vähitellen sukkulan keskitasoon ja muodostavat metafaasilevyn. Ne halkeavat pituussuunnassa kahdeksi samanlaiseksi tytärkromosomiksi. Jälkivaiheessa, anafaasissa, tytärkromosomit eroavat ja kulkevat sukkulan päihin. Loppuvaiheessa, telofaasissa, tumasukkula häviää, kromosomien kierteisyys purkautuu, ja ne häviävät näkyvistä. Kummankin kromosomiryhmän ympärille muodostuu tumakelmu, ja tumajyväset tulevat näkyviin. Tumasta on tullut kaksi identtistä tumaa. Mitoosin kestoaika vaihtelee 10 minuutista useihin tunteihin. Jakautumisten välisenä aikana, interfaasissa, kromosomit kaksinkertaistuvat seuraavaa jakautumisvaihetta varten.
b) meioosin eli kypsymisjakautumisen tuloksena syntyy lisääntymissoluja. Siihen kuuluu kaksi perättäistä tumanjakautumista. Ensimmäinen on vähennys- eli reduktiojakautuminen, jossa kromosomisto puolittuu. Toinen on tasaus- eli ekvaatiojakautuminen eli tavallinen mitoosi.